睡莲属植物细胞器RNA转录后加工研究取得进展(睡莲属植物细胞器RNA转录后加工研究取得进展)
植物细胞中有三个相对独立的基因组,即核基因组、叶绿体基因组和线粒体基因组,后两者常被称为细胞器基因组。RNA转录后加工,内含子剪接、RNA编辑、5’和3’端成熟等在植物细胞器基因组的基因表达和调控中很常见。植物细胞器RNA编辑已有报道,包括无油樟(Amborella trichopoda)及鹅掌楸(Liriodendron tulipifera)的细胞器RNA编辑,但内含子剪接及其与RNA编辑的相互作用研究较少。中国西南野生生物种质资源库利用PacBio Sequel平台的三代测序技术和Illumina Hiseq平台的二代测序技术,选择被子植物基部类群睡莲属植物品种黄乔伊(Nymphaea‘Joey Tomocik’)为研究对象,获取了其细胞器基因组和转录组序列;在此基础上,组装出完整的叶绿体基因组和线粒体基因组(图1),随后将全长转录组(Iso-seq)序列分别比对到两个细胞器基因组,并将去除核糖体RNA策略(rRNA-)建库(未经多聚腺苷酸富集polyA )的链特异性转录组测序(RNA-seq)数据(reads及Trinity组装转录本)分别比对到两个细胞器基因组,据此获取了睡莲属植物细胞器基因组在转录后RNA加工过程(内含子剪接及RNA编辑)的概貌。
研究基于全长转录组数据可以校正基于同源性的细胞器基因组注释结果。该研究发现,GenBank数据库中多数植物(包括拟南芥、水稻、无油樟等)的线粒体基因nad4-i2上下游的两个外显子间的边界存在注释错误。基于转录本比对结果,检测到睡莲属植物细胞器基因组中全部7个反式剪接内含子(叶绿体中的rps12-i1、nad1-i1、nad1-i3、nad1-i4、nad2-i2、nad5-i2、nad5-i3)的剪接证据,以及除转运RNA基因中的内含子以外的其它基因的全部顺式内含子的剪接证据。此外,该研究还首次检测到线粒体基因nad4(含三个顺式剪接内含子)的全部8种可能的内含子剪接产物;反式剪接和顺式剪接的发生互有先后,结果表明,细胞器基因组的内含子剪接是随机发生的,无先后顺序(图2)。
通过链特异性转录组测序数据直接比对后识别单核苷酸多态性(SNP calling),以及Trinity软件组装后的转录本比对相结合的方法,经筛选过滤,研究获得了睡莲属叶绿体基因组中98个、线粒体基因组中865个高可信的RNA编辑位点。比较发现,两种细胞器中RNA编辑绝大部分发生在编码区(其中以密码子第二位及第一位最多),80%以上的编辑位点编辑效率均超过0.6,非同义编辑前后氨基酸疏水性均增加,编辑位点上游-1位剪辑大多数为嘧啶(T和C),可以推断植物中两种细胞器基因组的RNA编辑可能有共同的起源和相同的机制(图3)。对比被子植物基部类群无油樟和木兰类鹅掌楸的细胞器基因组的RNA编辑位点,睡莲属的RNA编辑位点数目介于二者之间,三种植物叶绿体均为ndhD基因的编辑位点最多,线粒体均为nad4基因的编辑位点最多。序列比对后发现,除部分共有的编辑位点外,三个物种各有其特异性的编辑位点,由此可见,被子植物早期分支的RNA编辑位点丢失可能是物种分化后独立发生的。
细胞器基因组内含子剪接和RNA编辑的互作分析发现,RNA编辑在内含子和外显子区同时发生,内含子的RNA编辑对其自身的剪接十分重要。部分外显子中靠近内含子边界的RNA编辑位点则会受到影响,研究检测到内含子上游7个外显子和下游3个外显子中的RNA编辑位点有此现象(距内含子边界在2bp到39bp之间),须内含子剪接之后才可以被编辑,但部分外显子中靠近内含子边界的编辑位点不受内含子剪接与否的影响(如nad4 exon3中距离前后两个内含子28bp和27bp的编辑位点)。
相关研究成果以Organelle Genomes and Transcriptomes of Nymphaea Reveal the Interplay between Intron Splicing and RNA Editing为题,发表在International Journal of Molecular Sciences上。研究工作得到中科院重大科技基础设施开放研究项目的资助。

图1.睡莲品种黄乔伊线粒体基因组(含25kb重复)和叶绿体基因组的组装和注释

图2.睡莲属植物叶绿体基因组和线粒体基因组的顺式及反式内含子剪接
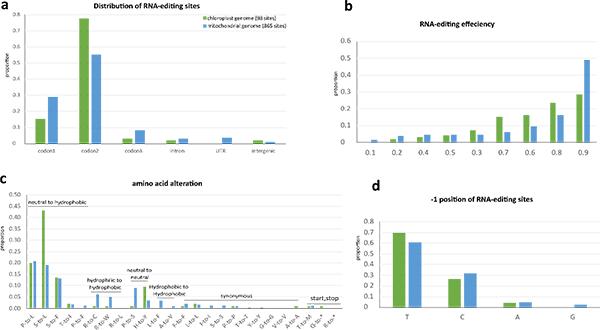
图3.睡莲属植物叶绿体基因组和线粒体基因组RNA编辑对比
来源:中国科学院昆明植物研究所
,免责声明:本文仅代表文章作者的个人观点,与本站无关。其原创性、真实性以及文中陈述文字和内容未经本站证实,对本文以及其中全部或者部分内容文字的真实性、完整性和原创性本站不作任何保证或承诺,请读者仅作参考,并自行核实相关内容。文章投诉邮箱:anhduc.ph@yahoo.com






