生石灰清塘的原理和方法(用了那么多年生石灰清塘)
提 要
本文报导了1981年6~7月对金州鱼种场的四个鱼池用生石灰排水和带水清塘后水质的变化情况。排水清塘池:pH 24小时可降至10以下,一周左右出现第二次高峰值(晴天可超过10)。在此pH高峰时溶氧也达最高值(有时超过20毫克/升);氨氮和活性磷酸盐有不同程度的增加,前者最高值4~5毫克/升,后者0.2毫克/升左右;其余碱度、硬度、亚硝酸盐、活性硅酸盐等亦有比较明显的变化;浮游植物4~7天达到高峰值(大于10毫克/升);轮虫5~7天出现高峰值(大于20毫克/升);枝角类的大量发生紧接轮虫高峰期之后;桡足类的成体生物量始终不大。带水清塘池的化学、生物变化程序和排水池相似,但在时间和量上有颇大差异。作者根据清塘后的水质变化规律结合鱼苗下塘问题进行了讨论。
生石灰清塘是我国淡水养鱼业中一种传统的清塘方式,其方法、机理和效果早就有过报导。倪达书等(1956)[1]曾用生石灰、巴豆、茶粕等药物进行清塘对比试验后指出,用生石灰清塘,杀灭野杂鱼类及浮游生物的能力较其他药物强得多,并且也能杀灭致病菌。陈觉民等(1975)也曾探讨过生石灰清塘的原理。
在国外,诸如美国、日本、苏联等也有过在水体中投放生石灰改良水质的报导。但是系统论述生石灰清塘后水质变化规律的材料,迄今尚未见到。
在生石灰清塘被普遍应用的我国池塘养鱼业中,弄清其水质变化规律,对池塘水质控制及鱼苗的适时下塘,都可提供必要的理论根据。为此,我们于1981年6~7月进行了这方面的试验研究。
# 一、池塘条件和试验方法 #
供试验用的鱼池是大连水产学院金州鱼种场的301、302、304、305号鱼池,面积分别为1.02、1.13、0.99、0.75亩。底泥较厚,平均在0.4米左右。有微渗漏现象,池水深度的变化是:301池由清塘后的45厘米增到60厘米;302池由43厘米增到60厘米;304池由4厘米增到46厘米;305池由8厘米增到76厘米。观测期间约有80%为晴天,6 ~7月平均水温为:301 池26.7°C;302池25.7℃;304池26.1℃;305 池24.4℃。
由于受沿海盐碱性土壤的影响,池水是氯化物类钠组Ⅲ型水,含盐量1.7~2.3g/L,氯离子为820~1200mg/L,硫酸根为150~240mg/L,总碱度为1.5~2.9me/L,钙离子为6.5~7.8me/L,镁离子为6.4~8.4me/L,钾钠离子合为340~580mg/L。
四个池塘中,301与302号池为带水清塘,304与305号池为排水清塘。清塘前人为搅起底泥。(305池排水前曾饲养鲤鱼)试验期间301与305池不曾施肥,302与304池先后施有机粪肥。各池清塘时的用灰量分别按亩施600、630、200、200斤计算,分别施入675、712、200、150斤。于1981年6月29日下午2点开始清塘,用水将生石灰调成浆状,然后均匀泼洒池内。以pH值普遍达到11以上为度。
观测的项目主要为水化学和浮游生物。同时也测定了水温和底栖生物。采样时用一个长把水舀,绕池一周均匀采取。除pH值和溶氧的水样外,其余水化学项目及浮游生物均用同一水样分别灌取。溶氧及pH值用釆样瓶分上、下层采样(304、305池前期因水浅而只取表层水样)。当时固定溶氧,然后带回室内测定。pH值马上带回实验室 用pHS-2型精密酸度计测定。并且,pH值和溶解氧始终分上、下午采样,而其他项 目,前三天分上、下午采样,之后于每天上午或隔天上午采样。采样后立即测定,在三小时内各项目全部测完。
浮游生物的浓缩及计数方法,按《内陆水域渔业自然资源调查试行规范》,底泥中的轮虫冬卵用底面积20~30平方厘米,长20~30厘米的圆筒型采泥器釆集3~5厘米厚的表层底泥,然后用食盐与蔗糖混合液使冬卵与泥沙分离。
化学分析的测定方法参照《淡水养殖水化学》(湛江水院主编,1980),其中活性硅的测定中,以高锰酸钾氧化掉还原性物质,以排除干扰。三氮水样用氢氧化钠和硫酸锌沉淀处理,其余水样均以4000转/分的速度离心处理10分钟。
# 二、实验结果 #
(一)水化学成分
1、pH值(图 1)
各池洒石灰清塘后,pH值迅速由清塘前的8~9上升为11以上。其后的变化,两组不同清塘方式的池塘各不相同。排水清塘池(简称排水池,下同),清塘后的第一天pH值即下降,第二天即可降到9以下。从第三天或第四天起,pH值开始出现清晨低下午高的波浪式变化。一般在清塘后的第七天前后会出现pH值的第二个高峰值,然后又下降,这个高峰值常常可达10以上。以后的pH值随各池塘条件而异。

带水清塘池(简称带水池,下同),清塘后第一天的pH值还有所上升,从第二天开始下 降。下降的速度比排水池慢,五 天后才降到10以下。10~11天后才开始出现上、下午的波浪变化,日变化幅度也小得多,清塘后第二个高峰值也小得多。

2、溶解氧(图2)
各池清塘后溶氧均下降。但其后的变化两组池塘相差颇大。排水池清塘后第一天溶氧有所上升,然后再下降。清塘3~4天后开始出现早、晩的波浪式变化,且日较差迅速增大。晴天下午溶氧最高值可达20毫克/升 以上,饱和度可达270%。
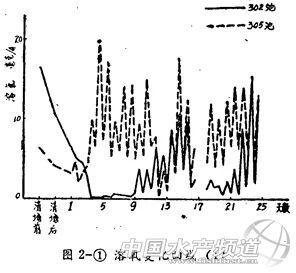
带水池溶氧在清塘后第三天降到最低值,以后连续数日保持在低水平。表底层溶氧在0.5毫克/升﹣以下的:301池有二天;施肥的302池有七天。第九天后才开始出现明显的早、晩波浪式变化,变幅比排水池小。

3、总碱度(图3)
各池清塘后碱度立即增加1~15 毫克当量/升。pH值高者碱度增加也多。清塘后碱度的变化,起初与 pH的变化方向相同。碱度下降情况依清塘方式而异:排水池从清塘后的第一天起碱度即下降;带水池清塘后 的一天多碱度还继续上升一些,然后再下降。

碱度的回升比pH早,排水池清塘1~2天后开始回升;带水池从第7~8天开始回升。碱度的第二个高峰值排水池在清塘后3~5天出现(2.8~2.9毫克当量/升);带水池高峰值不明显。
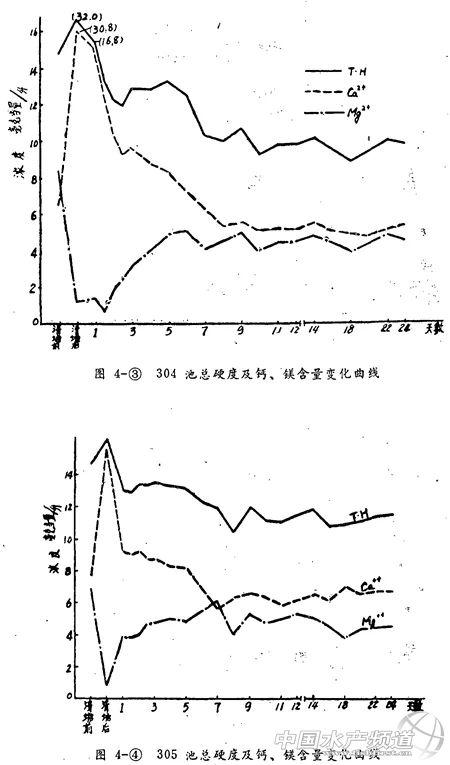
4、总硬度及钙、镁含量(图4)
各池清塘前水的总硬度在14~15毫克当量/升。钙、镁含量相近,甚至镁超过钙。清塘后总硬度增加,钙硬度也大幅度增加,略低于总硬度。镁硬度则大幅度下降,仅有0.6 ~1.2毫克当量/升。但两类池塘增减幅度有差别。
排水池总硬度增加幅度大,然后就迅速下降。其中钙硬度在下降,而镁硬度却在上升。七、八天后两项硬度趋于接近,并改变了清塘前的镁硬度高于钙硬度的状况,变为钙硬度高于镁硬度。
带水清塘后总硬度也有不同程度增加。pH值高者总硬度增加多,pH低者增加少。清塘后第一天硬度仍有较大的增加,这些均和碱度的情况一致。从清塘后第二天起,钙硬度和总硬度均下降。但下降速度比排水池慢。镁硬度在清塘半个月后才有较明显的增加。此时总硬度基本未变,钙硬度在下降。

5、铵盐、亚硝酸盐(图5)
清塘前铵盐含量,排水池的高而带水池的低。清塘后,无论施肥与否均大幅度增加。尤其排水池增加更快,未施肥的在清塘后二天达到最高值(3.88毫克氮/升),增加了2.6毫克氮/升。施肥的在清塘后三天达到最高值(4.99毫克/升),增加了2.8 毫克氮/升。以后均迅速下降。但施肥的下降较慢些。

带水池未施肥的铵盐含量上升较慢,到第11天才达高峰值(3.54毫克氮/升),増 加了3.4毫克氮/升施肥的上升较快,第三天达3.08毫克氮/升。然后下降,后又上升,在第九天达第二个高峰(3.52毫克氮/升)。

亚硝酸含量在清塘后均逐渐增加,施肥的增加得多。其中以带水池施肥的增加最多,最高达到0.38毫克氮/升。其他池的最高值仅在0.02-0.04毫克氮?/升之间。硝酸氮因测定方法存在问题,变化情况未搞清楚。
6、活性磷酸盐(图6)
各池清塘前的含量约在0.01~0.03毫克磷/升之间。清塘后立即降低到检测不出来。以后含量又增加,但数值波动很大,高者可达0.18-0.22毫克磷/升,一般在0.01~0.02毫克磷/升上下。

7、活性硅酸盐(图7)
各池清塘前含量均较高,约为12~13毫克SiO?/升。清塘后立即下降(排水池降为2.6~2.8毫克SiO?/升,带水池降为4.5~5.3毫克SiO?/升)。之后又回升,在 清塘后2~4天达一高峰值后又下降,以后的变化则依各池而异。

(二)生物状况
1、种、量变化
(1)各池清塘后浮游植物基本死亡,生物量不足1毫克/升(图8-①)但是其繁殖情况却依清塘方式而有显着差异:带水池的301、302池的浮游植物量是在清塘后第七天才开始明显上升的(大于3毫克/升),并且以后的增长始终很慢,生物量很少超过10毫克/升,平均只有2.9毫克/升和2.0毫克/升;而排水池的304、305池的浮游植物量在清塘后第四天就上升到3毫克/升以上,之后的发展虽有颇大的波动,但经常可以达到10毫克/升以上,平均为16.4毫克/升和26.3毫克/升。从生物量上看,清塘后浮游植物有两个高峰期:第一个高峰期是在轮虫大量发生前。这在排水池大约是清塘4-7天,带水池大约是清塘后10天左右;第二个高峰期是在轮虫、枝角类等滤食性浮游动物大量发生期之后,具体时间随池水中浮游动物量的多寡和延续时间的长短而不 同,通常都出现于生石灰清塘半月之后。

各池浮游植物的种类组成,在清塘前差异较大。包括绿球藻类、菱形藻(Nitzschia)、微囊藻(Microcystis)、壳虫藻(Trachelomonas)和兰隐藻(Chroomonas)等鞭毛藻和非鞭毛藻,而清塘后则有显着的共性,即初期为绿球藻类和小型菱形藻;中、后期分别出现隐藻(Cryptomonas)、衣藻(Chlamydomonas)、单鞭金藻(Chromulina)等鞭毛藻类。
(2)轮虫在清塘后4~6天开始出现,排水池的最初生物量较高(接近2毫克/升),而带水池只有少数个体出现。值得注意的是,两组池塘轮虫生物量的发展有极大的差异(图8-②)。排水池在轮虫出现后1~2天就达到20毫克/升以上的高峰期,而带水池却要再经过一周左右方能较大量的出现。有的(如301池)甚至始终不能达到上述的高峰期。
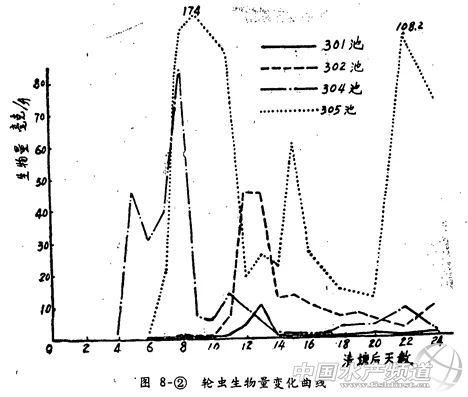
整个试验期间轮虫的平均生物量也是排水池高于带水池(前者约为后者的6倍)然而施有机肥的带水池302池的轮虫平均生物量,却和不施有机肥的排水池305池接近,都是每升十余毫克。
从生物量上看,无论排水池还是带水池,在清塘后一般都有一个轮虫的高峰期。在本试验的条件(水温25~26℃,底泥轮虫冬卵量100~200万粒/米2)下,排水池轮虫高峰期出现在清塘后5~7天;带水池轮虫高峰期出现在清塘后12~13天,大约相差近一周的时间。同时两组不同清塘方式的池塘,其轮虫生物量超过20毫克/升高峰期的天数也有颇大差别,其中排水池不施肥、不控制枝角类的305池为4天;排水池施肥并用敌百虫控制枝角类的304池为10天,带水池施有机肥的302池为2天;带水池不施肥的301池轮虫生物量最高只有10.72毫克/.升,始终没有出现上述高峰。
各池轮虫的种类组成十分近似。主要包括萼花臂尾轮虫(Brachionus Calyciflorus)、壶状端尾轮虫(B. urceus)、巨腕轮虫(Pedalia)和晶囊轮虫(Asplanchna),部分池塘还有少量的角突臂尾轮虫 (B. angularis)和多肢轮虫(Polyarthra)出现。
各种类的发生顺序多半是:萼花臂尾轮虫→壶状臂尾轮虫→巨腕轮虫→晶囊轮虫。角突臂尾轮虫和多肢轮虫等小型种类多半间插出现于上述种类之间,有时也能成为优势种。
(3)枝角类(图8-③)以体长1毫来左右的小型种类一裸腹水蚤(Moina)为主。于轮虫大量发生的时期开始出现。几个池塘生物量相差悬殊。304池因用敌百虫控制,平均生物量不足5毫克./升;施有机肥的302池裸腹水蚤生物量平均高达96.34毫克/.升;其余两个不施肥的池塘,平均生物量都不超过10毫克./升。

隆线水蚤(Daphnia carinata)在各池都有发生。此种大型种类多半出现在后期(清塘15天之后),其平均生物量不大(小于10毫克./升)。
2、发生程序(图9)
从图9-①-④清楚地看出:
(1)各池浮游生物发生顺序基本一致,最先出现藻类,然后是轮虫,紧接着为裸腹蚤、隆线蚤等枝角类,最后是桡足类的成体。

(2)各类浮游生物量达到高峰的时间,亦有一定的顺序。301、302、304池的轮虫高峰期都紧接浮游植物高峰期之后,并且浮游植物生物量又随轮虫生物量的急增而骤跌,此种因食物关系而呈明显反相关的现象在304池表现最为突出。305池上述顺序性不甚明显,但它在清塘后第五天出现的第一个轮虫高峰期仍然发生在第4天浮游植物大量繁殖(生物量5.2毫克./升)之后。


除用药物控制的304池外,轮虫高峰期之后,通常是枝角类高峰。此阶段浮游植物生物量不高(小于10毫克/升),只有待枝角类减少后(此时桡足类成体出现)其生物量才再次上升。

# 三、讨论 #
(一)水化学部分
1、pH值、碱度、硬度的变化
供试验的池水按阿列金分类法属于氯化物类钠组Ⅲ,水中钙、镁离子含量超过了HCO?-、CO?2-及SO?2-的总和。往这种水中泼洒石灰水,有上列沉淀反应发生:

这些反应的结果使水中的HCO?-及Mg2 被除去。部分Mg2 被Ca2 所替换,加上过剩的Ca(OH)?在水中的电离,致使水的pH升高,镁硬度减小、钙硬度增加、总硬度也增加。清塘后的碱度显然主要只能由氢氧根组成了。
清塘后,由于呼吸空气中的CO?及底泥释放CO?,可发生下列反应:
渗水带入的Ca(HCO?)?也可与Ca(OH)?反应:
这些反应的结果都是使水的pH值降低,总硬度、钙硬度和碱度均下峰。下列反应则使水中Mg2 含量增加,碱度和总硬度均上升,而钙含量则继续下降:
碱度升到高峰值时,pH值正在最低值。
当浮游植物大量发生后,光合作用要消耗大量的CO?,藻类会从HCO?-中夺取CO?,使生成CaCO?沉淀:
其结果是使碱度、硬度下降,pH值上升。植物光合作用的昼夜变化,使池水pH值出现昼夜变化。因为这时营养盐较丰富,藻类的敌害生物又没发展起来,遇到晴天,藻类繁殖很快,光合作用强烈,白天消耗的CO?在晩间得不到充分的补偿,则会造成pH值的极度升高,可能达到10以上(如304池)。据我们多次观察,对于排水池pH值的这个高峰值一般在清塘后的一周左右出现。具体日期及极值的大小与清塘后的天气有关。当轮虫大量出现以后,藻类被轮虫摄食,光合作用强度迅速减小,pH值又开始下降。
以上变化过程,对于排水池,由于石灰用量少,渗水量较大,CO?的补给速率相对较大,因而发展快。而带水池水量大,用石灰多,渗水量又少,CO?的补给速率相对较小,因而发展缓慢。
2、溶氧的变化
泼洒石灰后,因水中藻类被杀死,氧气只能从空气中补给,而这种补给速率一般是很慢的。水及底泥中有机物的分解使水中溶氧降低。排水池因水浅,当pH较高时,耗氧较慢,空气补给相对较大,溶氧会先上升一些后再下降,如304池清塘后第一天的情况。排水池pH下降快,浮游植物发生早,加上营养盐丰富,繁殖增长快,溶氧在清塘后2~4天即开始上升,并有早晚变化。其中施肥池溶氧上升较晚。
带水池pH下降慢、水深,空气补给显得相对小,致使池水较长时间处于缺氧状态。施肥池因耗氧大,缺氧状态持续时间就更长。
3、营养盐的变化
清塘后营养盐从总的来讲是增加的,氨氮的增加最明显。其增加的原因可能有两个方面:一方面是促进了底泥中有机氮的矿化及铵态氮的释放(钙离子的交换吸附作用);另一方面是被杀死的水生生物的分解矿化。这从溶氧与铵氮变化的关系中可以看出。溶氧下降或保持较低状态时(表明水中有机物分解较强烈),铵氮含量就增加。藻类繁殖起来后,铵氮含量就迅速下降。另外,铵氮下降时,亚硝酸氮含量都增加,表明硝化作用也消耗着铵氮,使其转变为硝酸态氮。遗憾的是由于测定方法问题,致使硝酸态氮的变化规律未测出。
活性磷和硅的含量在泼洒石灰后均立即下降。原因是它极易被氢氧化钙沉淀为 CaHPO?、CaSiO?等难溶物。胶态的硅和磷酸盐亦被吸附并沉淀。pH下降后,这些沉淀了的磷又会溶解,死亡的生物体中的磷也被释放,使水中磷含量增加。排水池可从渗水中得到磷和硅的补充。以后的变化则主要受到生物状况所左右。藻类多时,磷被消耗。硅藻大量繁殖可使硅含量大大下降。浮游动物滋生后,磷的含量又可上升。
(二)浮游生物的发生程序和轮虫高峰期
试验表明,清塘后各类浮游生物出现的时间和数量因清塘方式和施肥状况而异,然而各类浮游生物的发生程序却基本一致,即:藻类→轮虫→小型枝角类→大型枝角类 →桡足类。
如所周知,在养鱼池底泥中,广泛分布着各种藻类和浮游动物的休眠孢子、冬卵等,它们对药物有相当强的抵卸能力,生右灰清塘不仅对它们无妨碍,相反,还因为池中原有生物(包括敌害)被杀灭而创造了萌发后迅速增长的有利条件。因此,当清塘药力失效(排水池大约24小时,带水池需4~5天)后,各种休眠孢子和冬卵相继萌发。藻类孢子分布广泛,萌发速率也快,所以它出现最早。萌发后的藻类细胞在合适的池塘条件下,以分裂或孢子方式生殖,很快进入指数增长期,细胞数以每日几倍乃至几十倍的速度增长,其种群数量增加极为迅速[2]。
如排水池中304池,清塘后第四天的浮游植物量为4.82毫克/升,第七天就达51.8毫克/升,在有轮虫等滤食动物存在的情况下, 每天仍按2以上的倍数增加其生物量。绿球藻和菱形藻等小型单细胞藻类繁殖最快,所以它们首先大量发生。在藻类繁殖的同时,池底轮虫冬卵也开始萌发,据我们[3]近期的试验,轮虫冬卵在pH值11以下、水温20-25℃时,20小时左右就可萌发。排水池清塘后十几小时,pH值即可下降到11以下,所以在这类池塘中,清塘后1~2天就可以用密网捞到初萌发的小轮虫。而带水池却往往要等5-6天才有轮虫出现,这显然是pH的直接作用。由于冬卵密度的限制,直接从冬卵萌发出的第一代轮虫数是不多的,大约要经过4~5天的增殖,方可达到20毫克/升,加上清塘后有数小时高pH阶段和冬卵本身萌发所需时间,所以,即使在排水清塘的池塘也要一周左右池水中的轮虫才能达到高峰。304、305池清塘后只用5~7天轮虫就达到上述高峰,主要是试验期间水温比较高(平均26℃),另外,305池清塘前饲养鲤亲鱼,鲤鱼的活动对改善底泥中冬卵的分布很有帮助。加上底泥中轮虫的冬卵量(895万/米2)本来就比304池(225万/米2)为高,因此305池轮虫高峰期在清塘后5天就达到了,比条件与其相近似的304池提前两天。对于带水清塘的301、302池,由于pH值迟迟(4~5天)不能下降到11以下,加上清塘前未经拉网搅动底泥,所以尽管该两池底泥中的轮虫冬卵量并不少于304、305 池,它们的轮虫高峰期仍然推迟到了清塘后的第12~13天。
轮虫高峰期,浮游植物量显着下降,这显然是被食的结果。305池清塘后第七天,轮虫生物量剧增,而浮游植物生物量也急涨,这主要是肉食性晶囊轮虫占绝对优势。
池底淤泥中亦存在着相当数量的枝角类冬卵,它是池塘枝角类发生的主要根据。
据村上(1957、1959)[5]的材料,养鱼池中枝角类的冬卵分布极不均匀,在10厘米泥层中的量可达几万—几十万/米2,其量决定了春季池塘枝角类的发生量。金州鱼池常用敌百虫,枝角类及其冬卵量较少,另外,枝角类冬卵较轮虫卵萌发时需要更多的积热,萌发时间就长(25°C时需30余小时,较轮虫慢10多个小时),并且繁殖速度也不及轮虫。因此,清塘后枝角类的高峰就出现在轮虫之后。
桡足类的无节幼体大约与轮虫同时出现,但是由于它的世代周期长(至少七十二天),幼体必须几经蜕皮方可达到性成熟,故成体的出现,多半就在枝角类高峰之后了,有的池塘,甚至在整个鱼苗饲养期不出现大量的桡足类成体。
(三)从清塘后的水质变化规律看着鱼苗的适时下塘
本试验表明,生石灰清塘后,池水的化学成份和生物因子都同时发生着有规律的变化。此种规律在本试验的池塘条件(水温26°C、池底轮虫冬卵量200万/米2)下,排水池可模式化为:

从此模式可清楚看出,排水清塘后7~10天,无论从食物条件,还是水的化学成分看,都对鱼苗生长最为有利。因为此时正值鱼苗适口饵料——轮虫的高峰期;同时此阶段池水的pH值也降到9左右;溶氧在晴天中午可接近饱和,清晨也有3毫克/升以上,既不会使鱼苗得气泡病,也不会严重缺氧浮头,这时期的铵氮虽然还可能比较多(1毫克氮·升﹣1左右) ,但由于pH值较低,亦不会造成危害。可见,鱼苗在此轮虫高峰期下塘最合适,可谓“适时下塘”。从水质变化规律看,适时下塘的鱼苗,不仅在入池阶段生长迅速,而且以后生长也是良好的。实践表明,用生石灰清塘后,在一般池塘中自然繁殖起来的轮虫、枝角类等浮游动物,大约可供10万?亩﹣1鱼苗吃一周左右;而经人工控制(施肥、注水、控制敌害等)者,浮游动物大约能维持到鱼苗下塘后10~15天,其后由于鱼苗摄食量的急增,池中动物性饵料将供不应求,同时鱼苗还有可能受到浮游植物高峰期所造成的又一次高pH值的影响。所以,鱼苗还应当及时分塘。分塘时间以池 中浮游动物被食殆尽(大约在鱼苗下塘后10~15),鱼苗长成7~8分的乌仔头时为当,否则,鱼苗(特别是鳙、草鱼苗)生长将受到严重影响。
鱼苗在池塘轮虫高峰期下塘后的培育效果,近年来已为实践所证明。但是,在一些地区的鱼苗生产中仍流传着所谓“肥水下塘”和“清水下塘”的说法。先谈“肥水下塘",如果此肥水的概念是指池水浓绿(富含浮游植物,它既是池塘主要原初生产者,又可通过水色目测其肥度,应当是肥水的主要标志)的话,那么,在本试验的模式图中,就有可能是两个阶段:一是清塘后4~7天,轮虫即将达到高峰之前;另一是清塘15天之后。此两阶段的共同点是:pH值特别高(晴天最高值可超过10);溶氧高度饱和(有时达20毫克/升以上);浮游植物多而浮游动物少。显然,这时下塘,鱼苗缺少适口食物,还因溶氧过高而容易患泡病或直接受高pH值和氨态氮的危害。根据雷衍之等(1979)的试验,水温24~26.5℃、pH为9.5时、总氨(NH? NH? )对鳙鱼苗的24小时半致死浓度为1.1毫克/升;鲢鱼苗为0.55毫克/升。这个数值在清塘后4~7天的肥水阶段,是很容易达到的。因此而造成下塘鱼苗死亡的事故累见不鲜。所以,这样的肥水下塘弊病甚多,不应提倡。但是,浮游植物多的肥水是否一概不宜投放鱼苗呢?不是的。通常情况下,轮虫(主要是萼花臂尾轮虫等大型滤食性轮虫) 多的水是不会太肥的。可是有时候(多半在清塘后10来天,轮虫高峰期的未期),池水中存在着许多肉食性的晶囊轮虫或只能滤食小型藻类的龟甲轮虫(Keratella)、三肢轮 虫(Filinia)、多肢轮虫等小型轮虫时,鉴于它们对浮游植物、尤其是对较大型浮游植物影响不大,池水仍然可能比较肥,在这样的池水中产氧因子,耗氧因子都比较多,溶氧和pH值的变化多半为鱼苗所能适应,因此,这样的肥水还是可以投放鱼苗的。这恐怕也就是生产中有时候“肥水下塘”效果也不错的原因所在吧!但是,此种轮虫多的 “肥水”和前述很少轮虫的“肥水”从概念上容易混淆。所以,还是不提为好。
那么“清水下塘”又怎么样呢?当然,在本试验的模式表中,除开前述两个肥水阶段外,其余都是比较清瘦的了。刚刚清塘后2~3天的瘦水是根本谈不上放鱼苗的。在轮虫处于高峰期的阶段,浮游植物或多少,水或清或肥。可见,只有在枝角类大量发生的阶段,浮游植物最少,可视为典型的清水。在此阶段,一方面由于裸腹水蚤等枝角类的滋生,与之处于同一生态位的轮虫因滤食效率不及枝角类而造成种群的消退。枝角类几乎滤食掉全部浮游植物使水中溶氧极度紧张。此两点都不适宜于鱼苗的生长,这样的清水下塘自然是害多利少了。
总之,从清塘后水质变化的规律分析,无论“肥水下塘"还是“清水下塘”都只看到池水水质的表面现象,只有在轮虫高峰期下塘才抓住了鱼苗适口食物——轮虫这个关键,才算得上适时下塘。
以上是以排水池为例,论述了水质变化与鱼苗适时下塘的关系,至于带水池水质的变化,其基本程序和排水池一致,仅在变化时间上有很大差异,由于带水池亩用灰量大,池水渗入量又小,致使pH值长久不下降(5天还大于10),浮游生物迟迟不能发生。本试验的两个带水池都在清塘后10来天才开始出现轮虫,12天后轮虫数量才较大,并且由于带水池在清塘前难以充分搅动底泥,池底轮虫冬卵的分布得不到改善,所以轮虫冬卵萌发量少。虽然施肥可以促进其繁殖,如302池较不施肥的301池轮虫平均生物量高 7倍,并有两天达到20毫克/升以上的高峰期。但高峰期出现的时间已拖至清塘后近半个月,比排水池晚约一周,这是生产中不能允许的。加之以清塘时耗石灰量比排水池多出2倍以上,更为生产上所无法接受。可见,通常情况下,鱼苗池应采用生石灰排水清塘,不宜带水清塘。对于少数水源特别缺乏,或水中镁含量特高的池塘,采用带水清塘法可节约部分用水,还能减少镁的含量,有利于生物的发生。
# 四、小结 #
1、生石灰清塘后pH迅速上升到11以上,其后下降,排水池在24小时内可降至10 以下,4~7天时由于浮游植物光合作用而再度上升,晴天可达10以上;带水池5天后方可下降到10以下。随着pH的升降,池水碱度、硬度也发生相应的变化,特点是碱度的回升较pH为早,第二高峰值在清塘后3~5天,总硬度和钙硬度清塘后都有较明显的增加,而镁硬度则有所减少。
2、清塘后池水溶氧普遍下降,其后由于浮游植物光合作用而上升。排水池在4~7天时达最高值,晴天下午可达20毫克/升以上,以后随池水中生物的变化而有颇大的波动;带水池溶氧因浮游植物量少而变幅比较小。
3、铵盐、亚硝酸盐清塘后都有较大幅度的增加。前者在排水池最高值可达4~5毫克/升,带水池亦可达3毫克?升﹣1以上;后者最高值可达0.38毫克/升,一般不过0.03毫克/升左右。铵盐的含量随浮游植物繁殖而迅速下降。
4、活性磷酸盐和硅酸盐在清塘后立即下降,其后迅速回升,2~4天达高峰值。以后又因被浮游植物利用而减少,至浮游动物大量岀现后再度回升。
5、生石灰清塘后生物基本死亡,随着池水pH的下降,各类浮游生物相继滋生并达到高峰,在水温26℃的情况下,其基本顺序是:
清塘(4~7天)→浮游植物(10-15天)→轮虫(10-15天)→枝角类(15天后)→桡足类成体。此种顺序性是由各类生物自身的繁殖特点和食性所决定的。在人为干予下,可以改变其某一环节的数量及其存在时间,例如控制枝角类同时辅以施肥和注水,就可增加轮虫的生物量并可延续其高峰期的时间,从而有利于鱼苗的适时下塘。清塘后4~7天,当浮游植物第一次达到高峰时,因其强烈的光合作用,往往使pH极度上升至10以上,溶氧过饱和有时达20毫克/升以上,对鱼苗生长极为不利。生产中鱼苗下塘时务必避开这一 “危 险"时期。
参考文献
1.倪达书等,1956,生石灰、巴豆、茶粕清塘比较试验。水生生物学集刊,(1)
2.何志辉,1980,淡水生物学(下册),162页(讲义)。
3.李永函等,养鱼池轮虫休眠卵分布和萌发的研究(手稿)。
4.雷衍之、金送笛,1979,关于氨对鱼苗毒性的初步试验。辽宁淡水渔业,(3)
5.村上豊 1957。
作者:李永函 雷衍之 沈成钢
,免责声明:本文仅代表文章作者的个人观点,与本站无关。其原创性、真实性以及文中陈述文字和内容未经本站证实,对本文以及其中全部或者部分内容文字的真实性、完整性和原创性本站不作任何保证或承诺,请读者仅作参考,并自行核实相关内容。文章投诉邮箱:anhduc.ph@yahoo.com






