进化论和寒武纪生物大爆发(进化论不可撼动)

寒武纪发生于5.42亿年前【1】,现今大多数的后生动物门在寒武纪出现,似乎进化速度在寒武纪较快。然而寒武纪的持续时间却是有些难以界定和模糊的,靠谱的估计是持续了500万-1000万年【2】。然而,是不是寒武纪出现了现今大多数的后生动物门,就代表着现今所有的物种都出现在寒武纪呢?当然不是,比如哺乳动物、爬行动物、鸟类和昆虫等等这些常见的生物可没有出现在寒武纪。而且在寒武纪的鱼类(海口鱼)都和现代的鱼类有很大的不同【2】【3】。
寒武纪动物群中,最具有代表性的是尔吉斯页岩动物群和中国的澄江动物群。寒武纪很有可能经历过生物爆发事件,而由化石保存难易程度造成的假象的可能性并不大。

寒武纪时期的海口鱼
在解释寒武纪大爆发发生原因以前,有一点非常重要,就是寒武纪以前到底有什么?发生了什么?

我们先把目光放到十亿年前。在前寒武纪时的海洋中,生物的多样化主要体现在一些叫疑源类生物的化石上。疑源类出现在20亿年前左右,在持续10亿年后发生了一次辐射性的发展,具体表现在数量上、多样性上、体积上、形状的复杂程度上,特别是棘刺的体积和形状上。
有趣的是,它们在10亿年间发展的棘刺可能表明了防御掠食者的需要。新元古代的其他小有机生命群体也同样体现出了防备掠食者的防护特征【4】。而一些其他类别生物的寿命也支持了掠食者在当时带来的生存压力【5】。然而,通常在前寒武纪时期化石出现的非常缓慢。

陡山沱化石
在6.35亿年前的陡山沱中用显微镜可观测到的化石,或许代表着早期的两侧对称动物。一些学者认为这些化石是动物的胚胎和卵,而其中的一些化石或许是一些巨型细菌。同时一些学者指出,这些化石其实是真核生物的,而不是巨型细菌【9】。然而贵州小春虫,被认为是两侧对称的体腔动物【6】【7】,它或许也是仅仅被充入物质的气泡而已。【8】
这些化石表明了动物使用了最早的“快和硬”策略来对付掠食者的证据【10】【11】。

贵州小春虫
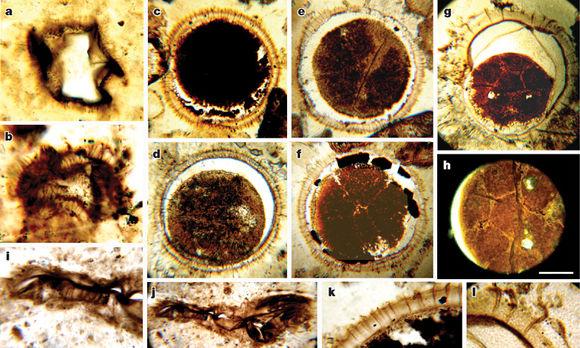
陡山沱中用显微镜可观测到的化石【9】
埃迪卡拉生物群
随陡山沱化石群后。接踵而至的就是埃迪卡拉生物群,大约从6.35亿年持续到5.42亿年左右【12】。在埃迪卡拉时期刚刚开始时,很多疑源类生物群在数亿年的时间里变化相对很小而且逐渐灭绝,而被一系列更新、更大并且时间短暂的多的物种所替代【13】。这次的辐射性演变,在化石记录中却是第一次,随后很快就出现了一大批人们不熟悉的、更大的化石群,而这些化石群被称为埃迪卡拉生物群【14】。并且这些生物繁衍了4千万年,直到寒武纪开始。大部分的埃迪卡拉生物群至少有数厘米长,并且明显比早期任何化石的体型都大。其中埃迪卡拉生物群中,有很多和以前与后来出现的生物非常不相似。有的像碟子、有的像袋子、有的像床垫。
然而,埃迪卡拉生物群中至少有些化石可能是寒武纪大爆发中一些重要的动物门的早期形态。比如早期的软体动物(金伯拉虫)【15】【16】,棘皮动物(Arkarua adami)【17】,节肢动物(斯普里格蠕虫)【18】。关于这些物种的争论仍然在持续着,因为埃迪卡拉生物群普遍缺乏更近代的一些生物的特征。然而,对金伯拉虫属于三胚胎层的两侧对称动物的怀疑至少很少【19】。这些生物对寒武纪大爆发的突发程度的争论非常重要。如果这些生物是现今动物门的早期成员,那会大大削减寒武纪大爆发的突发程度。

金伯拉虫化石
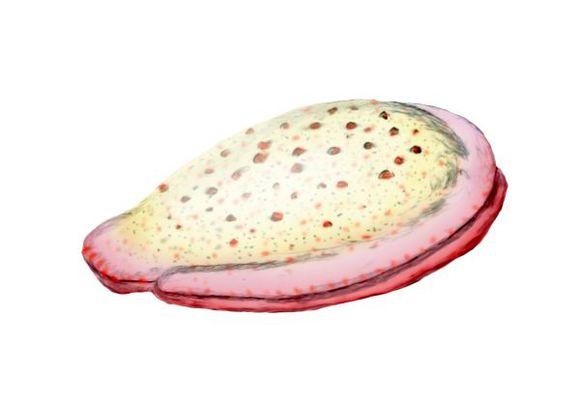
金伯拉虫复原
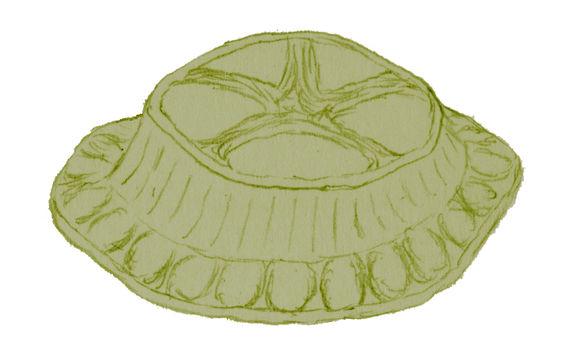
Arkarua adami

斯普里格蠕虫
埃迪卡拉晚期和寒武纪早期的一个有趣现象——进化军备竞赛
动物骨骼化的出现在埃迪卡拉晚期和寒武纪最早的时候,出现的第一种骨骼化的化石代表是管状虫以及一些带有疑问的海绵物种【20】。在埃迪卡拉快结束的时期(5.49亿年-5.42亿年前),科学家们找到了矿化后的克劳德管,Namacalathus,以及其他十多个有机体的化石,以及其他管状类的生物。
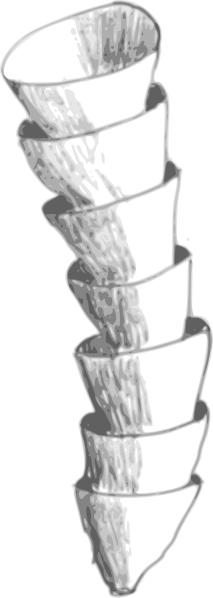
克劳德管复原图

Namacalathus复原图
埃迪卡拉的管状类生物通常存于叠层礁石的碳酸盐中等,这表明它们可以存活于对其他动物都很不利的环境中【21】【22】。
尽管这些化石也同样难以归类,但是它们在两个方面很重要。首先,这些生命是最早利用碳酸钙来为自己建造硬壳的【23】。其次,这类管状生物利用这些硬壳可以不同程度的防御掠食者和相对恶劣的生存环境。比如,克劳德管上的洞口则可能是被一个足以咬穿硬壳的掠食者攻击过的一个证据【24】。 这表明掠食者和它们的猎物在这个时期可能存在过激烈的“进化军备竞赛”。而进化军备竞赛假说是解释寒武纪大爆发的一个可能性之一。
所以,寒武纪大爆发的剧烈程度到底有多大?
寒武纪大爆发是困扰达尔文的一个主要难题,他认为这可能是反驳他的自然选择理论的一个证据。由于当时的在寒武纪以前的化石证据稀少,所以要解释寒武纪大爆发格外困难。
然而,如今随着寒武纪以前的化石证据的增多。我们对寒武纪大爆发的了解也就越多。如果埃迪卡拉的金伯拉虫是类软体中的原肢类动物,则表明原肢类和后口动物世系早在5.5亿年前就分化了【25】。就算它不是原肢类动物,它也被广泛的接受为两侧对称动物【25】。而且看起来较为现代的刺胞动物在陡山沱找到,这个表明刺胞动物和两侧对称动物早在5.8亿年前就已经分道扬镳了【25】。
在寒武纪以前出现的动物群,某种程度上削弱了“大爆发”这个词汇。它们不仅显示了动物的出现仍然是渐进的趋势,并且它们的多样性进化的速度可能并不如原先认为的那样快。实际上,统计学分析上显示,寒武纪大爆发不比生命历史上的其他辐射状演变的事件速度更快【26】。然而,寒武纪的爆发确实演变出了一些具有革新意义的特征,比如抵抗装甲,只在动物进化历史中发生了一次。
同时,比较传统的一个观点:“所有的门诞生在寒武纪”,现在来看是有错误的。分子生物学显示至少有六种动物的门处在寒武纪以前。同时,所有的植物出生在寒武纪以后。而目前的证据表明只有11/32的多细胞动物门出现在寒武纪,有8种处在寒武纪以后,一种处在寒武纪以前。剩余的化石未找到【27】。

尽管门在寒武纪发生了多样性的演变,很多门中的关键生物群体的典型代表都没有出现在寒武纪,而是在显生宙的后期才出现【28】。
那么,造成寒武纪和埃迪卡拉生物群的生物“大爆发”和多样化的原因是什么?
这些可能性的解释主要分为三大类:环境上的,发育上的和生态上的改变。
环境的改变
1.氧气含量的增多:
地球最早期的大气是没有氧气的。氧气的产生是由最初的光合作用,比如蓝藻。最初大气中的氧气含量并没有出现显著增加的趋势【29】。它们很快与石头周围和海洋中的铁和矿物质发生氧化还原反应。当这些反应达到一个饱和点时,氧便能以O2形式继续存在。随后大气中氧气的含量在25亿年前才开始逐渐增多。
从下图中的随地质学时间大气中氧气的分压可以直接看出氧气的含量。在38亿5千万前到24亿5千万年前,大气氧气含量基本为零。24亿5千万年前到18亿五千万年前,氧气含量显著增加。18亿五千万年到8亿五前万年时,氧气含量维持在稳定状态。8亿5千万年前到5亿四前万年前时,氧气含量开始持续上升。而到5亿4前万年时,氧气的含量达到了一个峰值。而这个时候恰恰是寒武纪所处的年代。
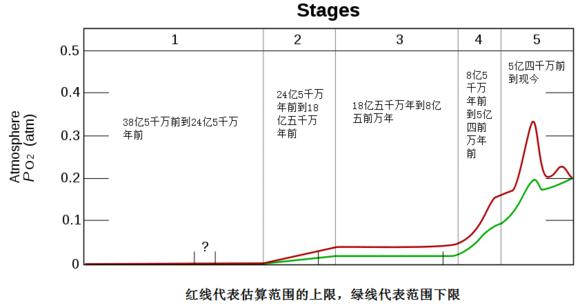
【30】
并且,早在寒武纪以前,就体现过氧含量与真核生物的多样性呈正比的关系【31】。所有现存真核生物的最后共同祖先生活在18亿年前。而在8亿年前,化石记录中出现了明显的真核生物复杂性和数量的增加【31】。
在真核生物的多样性发生以前,科学家们认为它们生活在一个含硫量高的环境。硫化物会阻碍好氧生物的线粒体功能,限制可以驱动新陈代谢的氧气含量。而海洋中的硫化物在8亿年前的含量开始减少,这进一步支持了氧气在真核生物多样性上的重要性【31】。
还有:在5.8亿年前的噶斯奇厄斯冰期前和期间,深海沉积物中铁的含量表明,当时的深海环境是缺氧的,然后才开始变得含氧。在噶斯奇厄斯冰期后,已知最早的埃迪卡拉生物群的成员就马上崛起。这个支持它们的进化和氧含量之间的关系【29】。
氧气的缺乏可能防止更大更复杂的生物的崛起,并且一些新陈代谢在缺少氧气时会被抑制。不过在显生宙某一时期,发生了同样的缺氧海洋环境,但是那里的动物并没有受到影响。这说明进化和氧含量之间的联系并不是足以令人信服和完全肯定的。所以,氧气成为生命发展为复杂形势的必要性,不比液态水和初级生产力更强【32】。当然,这并不能排除氧气含量对寒武纪生物群的影响。
2.雪球地球:
在新元古代晚期(持续到埃迪卡拉时期早期),地球一直在被巨大的冰川作用所影响,大部分表面积都覆盖着冰层。这会导致巨大的生物灭绝事件,造成种群遗传瓶颈效应。然而,当时陆地上的火山喷发时放出的CO2浓度是现今水平的350倍,在造成了剧烈的温室效应的同时,也令雪球地球迅速结束。
从一个雪球地球的结束到紧接着的极端温室效应,会让新元古代生命的进化历程经历一个强烈的选择压力。很多统治新元古代的原核生物从这场环境的剧变存活下来,并且没有受到影响。而很多真核生物的门在晚期新元古代前都发生了进化现象,说明它们都从强烈的选择压力存活了下来。然而,一场雪球地球带来的巨大冰川化会令当时变成的环境变为“过滤器”,造成了一系列的“种群遗传瓶颈效应”,随之而来剧烈的温室气候造成了“大坝开闸大放水”的效应,在这两个截然相反的效应的互相作用下,这可能导致了雪球地球结束后带来的埃迪卡拉生物群多样化辐射状演变。【33】
然而,雪球地球事件早在寒武纪开始以前就发生了,所以要想搞明白即使是一系列的瓶颈效应是如何能造成这么多的多样性不是一件容易的事情。冰冷的时期可能拖慢了体积比较大的有机体的演化。【34】
3.寒武纪海洋中钙浓度的改变:
新的研究表明,火山极其活跃的中央海脊会令海洋中的钙浓度变得非常巨大,可以为海洋生命提供建造骨骼和坚硬身体部分的原材料。并且研究者通过钙对海绵的细胞粘附分子作用来表明钙浓度对寒武纪多细胞生物所扮演的角色,为什么会导致多细胞的后生动物大爆发。
海绵是目前已知最原始的后生动物,并且在寒武纪以前就已经出现。所以海绵是个不错的研究的起点。
首先,同种识别能力可能是用来防止嵌合现象带来的负面效应,比如生殖细胞的寄生等。而海绵的幼虫在习惯上会融合在一起而形成可遗传的杂交成体。然而成体海绵却拥有精准的同种识别能力(注意,海绵可不是一个“种”),可以很好的防止不同种类个体互相杂交【35】。
下图就是海绵中物种和物种之间的细胞粘附,这个过程基于海绵蛋白聚糖的钙相关碳水化合物-碳水化合物相互作用。
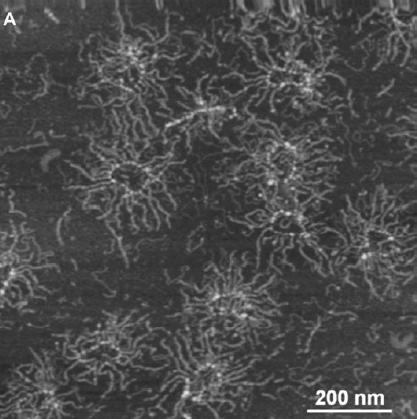
而在寒武纪海水中钙的浓度开始增加时,当时已经存在可以同种识别的原始后生动物。而这些原始的后生动物所携带的细胞粘附分子会随着钙离子浓度的增加而变得更稳定,需要更长的时间被分解。只有当钙的浓度增加时,才可以令细胞间的多价染色体黏合更加强壮,可以令个体表面束缚的后生动物--如海绵,保持完整并直到它们生殖。而不会被很快的拆散掉。这个情况可以令基因组成维持在多细胞生物的个体上,并且可以被持续的遗传下去。寒武纪时期的拥有同种识别能力的原始生物遇上了钙离子浓度增大的时机,而令它们的细胞粘附分子强度随着钙浓度增大而增强,大大的提高了诸如海绵这样的无柄后生动物的生存能力,这有可能最终导致了寒武纪大爆发【35】。
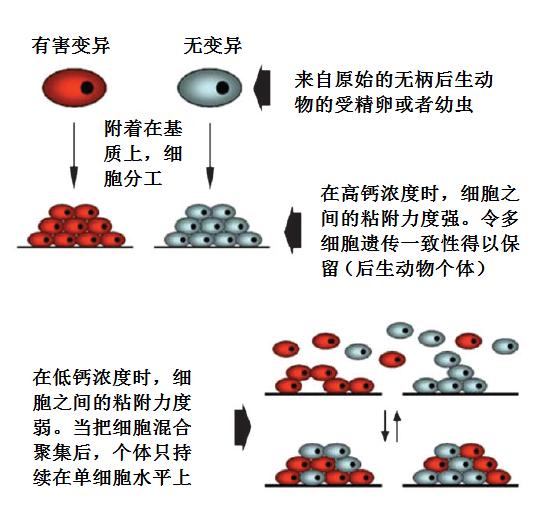
下图用卡通图像总结了高和低钙浓度对拥有钙依赖性细胞粘附分子的原始无柄后生动物的效果。早期能识别异己的后生动物能够发展处基于碳水化合物的细胞粘附能力,其在前寒武纪时的低钙浓度环境下寿命太短,难以保持在一起足够长的时间,无法令后生动物个体稳定持续下去。而在寒武纪时,钙浓度增高的情况下,便能更长时间更稳定的保持在一起,得以维持下去。【35】
同时高钙浓度有利于贝壳等的生成,为生物带来更多的生存优势【35】。
4.大不整合面引起的寒武纪大爆发
大不整合面是指的在地层中,有很大年代、起源和结构差别的沉积岩和沉积岩之间,或者沉积岩与晶层之间的接触面。这些巨大的差别象征着地质年代上足以产生一群山然后全都被侵蚀掉的时间。【36】
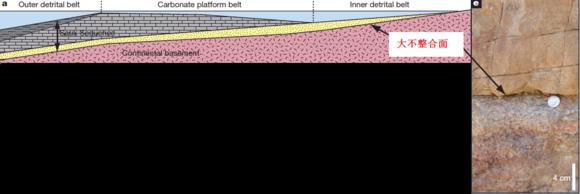
新的地层学和地球化学的证据表明,在5.4亿年到4.8亿年前的早期古生代的海洋沉积物中记录着浅陆缘海的扩张现象和异常形式的化学沉积物,这表明海洋碱性的增加和化学风化作用的变强。这些现象是由新元古代时期广泛分布的大陆长时间被侵蚀造成。而随后就伴随着寒武纪以后的显生宙第一次大陆范围的海侵现象时留下的风化层和基岩,其结果就是产生了大不整合面【37】。
除了在第3条说明的原因产生的钙离子以外,同时另外一种可以提供大量的离子的来源则可能是广泛的腐蚀结果。由这个结果造就的大不整合面可以带来巨量的离子,这令生物矿化提供了难以想象多的来源,可以令生物形成贝壳或者骨骼,而大不整合面为生物矿化的进化提供了“导火索”,进而引发了寒武纪大爆发【37】。
发育方面的解释
有一系列的解释寒武纪大爆发的理论是基于一个概念,这个概念是动物从胚胎到成体的发育时,微小的改变就可以导致最后成体非常大的改变。比如Hox基因,控制着一个胚胎的多器官单区域到底会发育出哪个器官。举个例子,如果一个特定的Hox基因被表达,那个区域会发育出一条腿。而如果另一个不同的Hox基因在那个区域被表达时(一个微小改变),反而会长出一个眼睛(巨大改变)。
这种体系需要一个条件,那就是大范围的不一致性可以从少量基因中出现。然而,将这个条件与大爆发联系起来的理论需要面对这样一个难题,就是为什么这种发育体系的起源可以自动导致多样性和不一致性的增加。不过,前寒武的后生动物的分子数据表明,很多在寒武纪已经完善建立的遗传结构确实可以的在大爆发中扮演一个角色,比如Hox基因。【38】
生态方面的解释
1.埃迪卡拉末期的大灭绝:
大灭绝的证据包括埃迪卡拉生物群化石记录的消失,还有一些贝壳类化石如克劳德管的消失,以及还有δ13C记录中的扰乱。【39】【40】
生物大灭绝后,往往会伴随着适应辐射状演变。因为幸存的生命会进一步扩张和利用大灭绝造成的生态位的空缺(比如恐龙灭绝后,哺乳动物的崛起以及多样化的产生)。
2.装备进化竞赛

躲避或从掠食恢复的能力往往决定着生死,因此捕食和被捕食是自然选择中最为强烈的一个成分。猎物面对的适应压力,相比掠食者要更大。如果掠食者失败,它丢掉的是一碗饭。而如果猎物失败,它丢掉的是生命【41】。二者在互相拼装备时,随之而来就是选择压,竞争越激烈造就的选择压就越强烈。而强烈的选择压则可能会造就进化的加快以及辐射状演变。寒武纪时期的那些可以破坏动物贝壳的大型掠食者,一直被认为是选择压的来源,并且对寒武纪带有硬骨骼和贝壳类动物的大爆发有着不小的影响【42】。
在埃迪卡拉纪和寒武纪过渡时期,发现了不少被击穿并咬破的骨骼或者贝壳化石。最引人注目的一个化石证据是,一个被掠食者攻击过的早期寒武纪纯真滇东贝的贝壳。其被保存的非常完好,甚至保留下来了一条长长的肉茎。同时贝壳上面的咬痕也清晰可见,而且还有康复时的痕迹【42】。
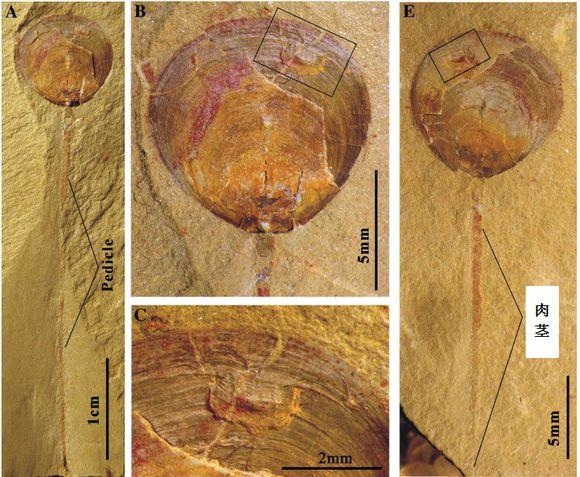
纯真滇东贝康复后的贝壳都会留下拉长的肉茎,这表明了它们从掠食者的攻击都存活下来,并且保持在受到攻击时的原处。发现了这些没有被攻击致死的贝壳,表明腕足动物面临着掠食者带来的选择压,自然选择会强硬的挑选那些出贝壳更坚固耐用的家伙们【42】。
然而,有证据表明早在寒武纪前,掠食者就广泛存在。比如,前文提到的疑源类增强的棘刺,还有克劳德管上被击穿的洞口。所以,尽管装备进化竞赛可能会剧烈影响大爆炸时产生的生物身体形态,对生物爆发进一步推波助澜。但掠食者的出现不太可能是寒武纪大爆发的导火索,而更有可能是上文提到的矿物质改变和大不整合面的产生。然而,掠食者的确在寒武纪时明显变得强大,比如发展出了击穿贝壳的策略。【42】
3.浮游生物的体积和多样化的增多
地球化学证据表明:自早期原生代开始,浮游生物的总质量和现今的水平相似。在寒武纪开始以前,它们的尸体和残屑过小,难以快速的掉在海床上。这表明它们的尸体在沉入海底前会被一些食腐动物和化学步骤破坏掉。
中型浮游动物是浮游动物中个体比较大的一类。而早期寒武纪的物种会从海水中过滤进食这些显微镜可见的微生物。这些中型浮游生物体积更大,因此其残渣和尸体会沉淀的相对更快。这会给中层和底部的海水区域提供新的能量来源和营养物质,给生命开启众多的生活方式。如果这些浮游生物的尸骸没有被吃而沉积到海底,它们会被逐渐埋葬,将一些碳从生态循环中带走,而增加海水中氧气的浓度。【43】
最初的食草中型浮游生物可能是海底动物的幼虫。这种成体和幼体生活在完全不同的领域的繁殖方式,很可能是在埃迪卡拉纪时期海底掠食者逐渐猖狂,而选择的一种进化策略。【44】【45】
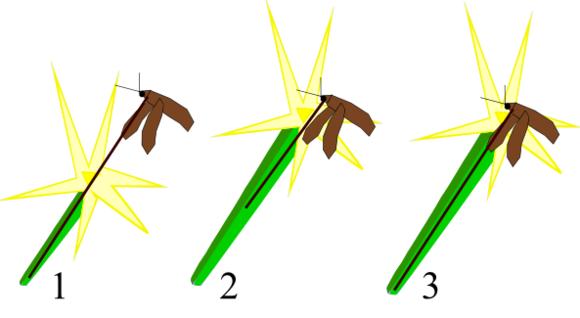
其实,这些中型浮游生物引起的就是共同进化作用。后生动物通过共同进化作用,可以增加不少多样性的演变【46】。这意味着一个生命的特征会导致另一个生命的特征随着发生进化。不同物种之间的相互作用会令它们共同发展。比如,掠食者的进化会导致猎物发展出更多的防御策略,或者发展出更多的逃跑策略。而为了应对猎物的策略,会迫使掠食者分化为两个不同的物种来应对猎物。一个擅长追踪猎物,而另一个擅长打破猎物的防备。实际上,共同进化是非常精妙和敏感的。然而,照这样的方式进行下去,确实可以造成多样性发展。四分之三的现存物种都是动物,而剩余的大部分物种都是和动物共同进化形成的。【46】
下图展示了共同进化的一个形象的例子:
4.对生态系统的改造
进化出的新物种会不可避免的改造周围的环境。比如在泥盆纪时期,陆生动物的出现和对陆地的侵略,对全球性的沉积物循环和海洋营养物质造成了影响,而这个事件可能和泥盆纪物种大灭绝有关。而类似的事件在海洋里面也发生了,只是尺度比较小。比如海绵会过滤一些微粒,并把它们以更容易消化的形式存储在泥土中。而会挖掘的动物也能令一些其他动物无法得到的资源变得可以获得。这些行为都会对生态系统进行改造。
总结
如果我们总结一下所有的原因,会发现寒武纪会出现生物大爆发的现象并不出乎意料。如果我们算笔账:
寒武纪的氧含量大大增多,这对一些生物的生存变得有利。
寒武纪以前,雪球地球就已经结束,然后就伴随着剧烈的温室效应。“瓶颈和大放水”的互相作用,虽然使埃迪卡拉生物群面临了强大的选择压。但是无疑给随后的寒武纪铺好了发展的温床。
此时,寒武纪前的埃迪卡拉末期,生物群面临大灭绝,而它们的大灭绝,正好空出了大量的生态位。而幸存的物种可以趁机崛起,在寒武纪占领这些生态位,大肆的发展。
寒武纪时大不整合面的产生和火山运动造成大量的矿物质离子涌出,给生命发展骨骼和贝壳等提供了非常好的机会和来源。而发展出贝壳等的一系列创新行为,更会导致以此为基础的进化多样性尝试。
贝壳、坚硬的身体部分等随后会更进一步激发掠食者和猎物之间激烈的“装备进化竞赛”,而这很有可能对生命大爆发的发展进一步推波助澜。
激烈的进化竞赛,又会进一步导致共同进化,推动生物改早环境。而这些因素可能还继续互相影响。
从这里,我们似乎可以看到:当生命的发展渡过一个艰难的瓶颈,突破那个关口后,只要努力打开一扇门,随后就会发生连锁反应般的继续打开其他的门,因为这些门都是互相影响着的。而寒武纪的生物也可以说是非常的幸运,很多好事都被它们赶上了。那么,我们或许应该期待在寒武纪的时候,生物会发生一次大爆发现象。生命自会寻找出路,当大自然已经在寒武纪把路很好的铺在生命面前,那么我们为什么不期待生命会顿时崛起前进呢?
引用的文献
Ref:
1.en.wikipedia.org/wiki/Cambrian_explosion
2.talkorigins.org/indexcc/CC/CC300.html
3.en.wikipedia.org/wiki/Haikouichthys
4.nrm.se/download/18.4e32c81078a8d9249800021552/Bengtson2002predation.pdf
5.paleobiol.geoscienceworld.org/content/34/1/1.extract
6.en.wikipedia.org/wiki/Vernanimalcula
7.ncbi.nlm.nih.gov/pubmed/15178752
8.sciencemag.org/content/306/5700/1291.1.full
9.nature.com/nature/journal/v446/n7136/fig_tab/nature05682_F1.html
10.nature.com/nature/journal/v391/n6667/full/391553a0.html
11.nature.com/nature/journal/v442/n7103/full/nature04890.html
12.en.wikipedia.org/wiki/Ediacaran_period
13.onlinelibrary.wiley.com/doi/10.1111/j.1475-4983.2006.00613.x/full
14.adsabs.harvard.edu/abs/2008Sci...319...81S
15.nature.com/nature/journal/v388/n6645/abs/388868a0.html
16.sciencemag.org/content/288/5467/841.abstract
17.az.oxfordjournals.org/content/38/6/965
18.gsa.confex.com/gsa/2003AM/finalprogram/abstract_62056.htm
19.onlinelibrary.wiley.com/doi/10.1002/bies.20507/abstract
20.sciencemag.org/content/279/5352/879
21.geology.geoscienceworld.org/content/29/12/1091.abstract
22.paleobiol.geoscienceworld.org/content/26/3/334.abstract
23.paleobiol.geoscienceworld.org/content/26/3/334.abstract
24.palaios.sepmonline.org/content/18/4-5/454
25.dev.biologists.org/content/129/13/3021.full
26.ncbi.nlm.nih.gov/pubmed/21680426
27.talkorigins.org/indexcc/CC/CC300.html
28.journals.cambridge.org/action/displayAbstract?fromPage=online&aid=623&fileId=S000632310000548X
29.ncbi.nlm.nih.gov/pubmed/17158290
30.en.wikipedia.org/wiki/History_of_Earth#/media/File:Oxygenation-atm.svg
31.ncbi.nlm.nih.gov/pmc/articles/PMC3158185/
32.onlinelibrary.wiley.com/doi/10.1111/j.1472-4669.2009.00188.x/abstract
33.sciencemag.org/content/281/5381/1342.long
34.nrm.se/download/18.4e32c81078a8d9249800021552/Bengtson2002predation.pdf
35.mbe.oxfordjournals.org/content/26/11/2551.full.pdf html?sid=d236f567-eab0-427b-96bc-40d7bad8dbd7
36.en.wikipedia.org/wiki/Great_Unconformity
37.nature.com/nature/journal/v484/n7394/pdf/nature10969.pdf
38.nature.com/nature/journal/v399/n6738/full/399772a0.html
39.sciencedirect.com/science/article/pii/S1871174X06000485
40.sciencedirect.com/science/article/pii/S0016703706006569
41.ncbi.nlm.nih.gov/pubmed/42057
42.sciencedirect.com/science/article/pii/S0031018211000137
43.Ecology and evolution of Cambrian plankton
44.paleobiol.geoscienceworld.org/content/31/2_Suppl/36.abstract
45. The Ecology of the Cambrian Radiation
46.onlinelibrary.wiley.com/doi/10.1111/j.1475-4983.2006.00613.x/full
文/怀疑探索者、量子幽灵
,免责声明:本文仅代表文章作者的个人观点,与本站无关。其原创性、真实性以及文中陈述文字和内容未经本站证实,对本文以及其中全部或者部分内容文字的真实性、完整性和原创性本站不作任何保证或承诺,请读者仅作参考,并自行核实相关内容。文章投诉邮箱:anhduc.ph@yahoo.com






